Senita Cactus and Senita Moth Mutualism

During 1995 and 1996 field studies in the Sonoran Desert, Dr. T.H. Fleming (Professor Emeritus, University of Miami) and I discovered the third known pollinating seed-consuming mutualism involving the behavioral adaptation of active pollination and a morphological adaptation for pollen collection and dispersal. The senita cactus/moth mutualism parallels the two classic cases of coevolved mutualisms between yucca/yucca moths and fig/fig wasps. Other pollinating seed-eating interactions include globeflower/flies, Lithophragma/Greya moth, Silene/Hadena moth, and Glochidion/Epicephala moth interactions, but only senita, yucca, and fig systems are known to involve such behavioral and morphological adaptations. Senita cacti benefit from pollination, but incur costs due to larval fruit consumption. Moth larvae benefit from fruit food resources, but incur costs to larval survival from fruit abortions.
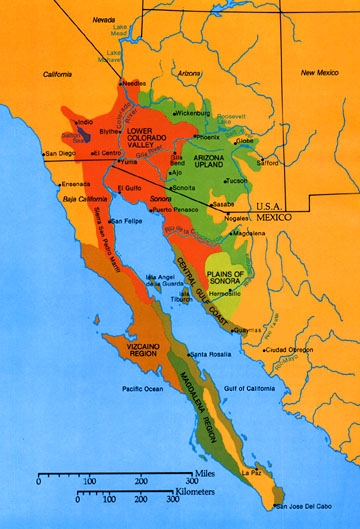
Both senita cacti and senita moths are endemic to the Sonoran Desert of North America. They are distributed throughout Baja California and Sonora in Mexico, but within the United States they only occur in Organ Pipe Cactus National Monument in Arizona. The two species of senita cacti, Pachycereus schottii and P. gatesii, were formerly Lophocereus schottii and L. gatesii, but recent phylogenetic analyses suggest returning them along with their closest relative Pachycereus marginatus to the Lophocereus genus. Only one locale of P. gatesii occurs, which is at the southern tip of Baja California.

Senita is a long-lived (>75 years) columnar cactus that can produce 100s of 2-5 m stems radiating from its base. Upon reaching reproductive maturity, senita flower every year during spring and summer months. Ribs along stems bear many areoles (spine bearing reproductive pads) that can produce one or more flowers per night. Senita can produce upwards of 3,000 flowers during 3-5 month flowering season. Flowers open and anthers dehisce at sunset and then close within 12 hours, but typically prior to sunrise. Senita are entirely self-incompatible, obligate outcrossers. Hermaphroditic flowers have an inferior ovary, one pistil, 100s of ovules, and 100s of anthers. Only 50% of flowers produce any nectar at all, with an average 0.5 ml in nectar producing flowers. Mature fruit are produced within 30 days containing ~160 seeds per fruit with >95% viability.

The senita moth, Upiga virescens (Cambidae), is a small moth with forewings 7-10 mm in length. It is the only species in its genus. Until recently, the only report of U. virescens was its species account. All life stages of U. virescens are associated only with senita cacti. Larval growth conforms to Dyar's rule, but unlike other Lepidoptera, only three larval instars have been identified. Adult moths mate on spines, female moths oviposit in flowers, larvae consume immature fruit, and pupation occurs in cactus stems.
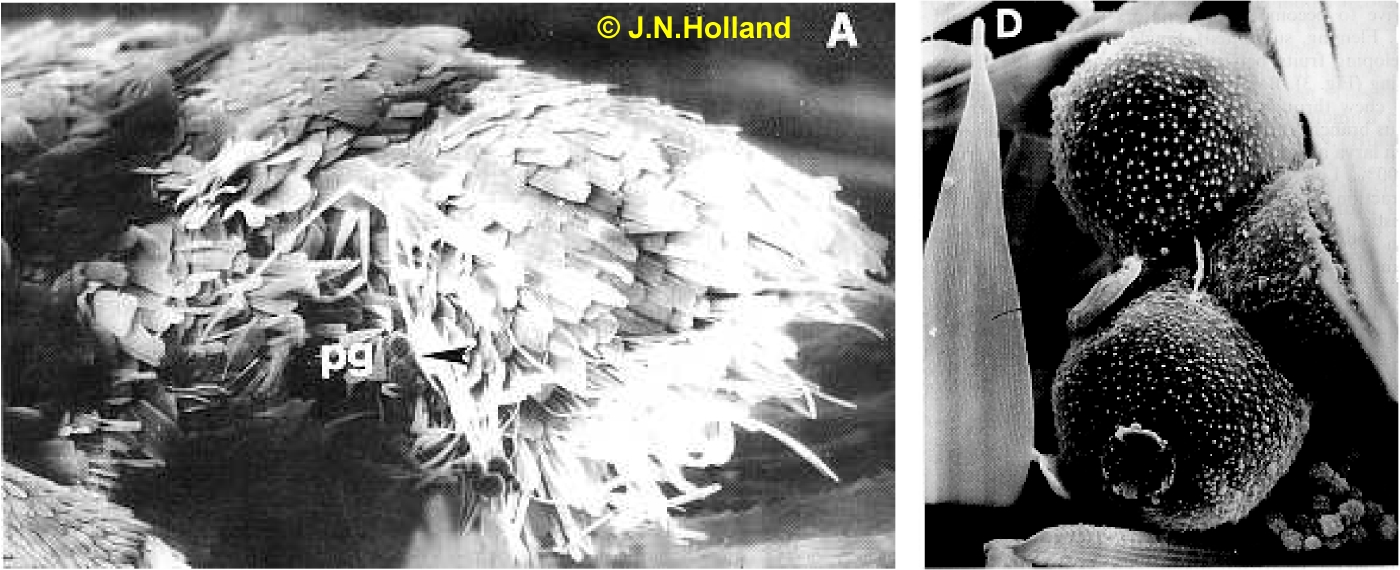
Senita moths are commonly seen mating on spines of senita cacti. Both male and female moths visit flowers for nectar, though most often flowers contain little to no nectar. Unlike male moths, however, female moths have an elaborate and consistent set of behaviors they exhibit during flower visitation. Immediately following sunset, female moths actively collect pollen from flowers using their pollen brush, presumably a morphological adaptation which consists of elongated scales on the ventral side of female abdomens. The pollen brush facilitates the collection, adhesion, and transport of pollen grains (pg in adjacent picture) among flowers. After collecting pollen, moths depart that flower and fly to another plant, as senita cacti are entirely self-incompatible.

Upon arriving at a flower that has not already been oviposited, as female moths avoid such flowers, female moths first exhibit a presumably adaptive behavior which has been named active pollination for yucca moths and fig wasps. For senita moths, this first behavior of flower visitation consists of climbing onto the flower's stigma in a head-down position and actively rubbing her abdomen onto the lobed stigma, thereby transferring pollen grains (~2,600) from her abdomenal pollen brush to the stigma. In doing so, senita moths actively pollinate flowers, rather than passively pollinating them by incidentally bumping into stigmas and transferring pollen while obtaining a food resource (nectar, pollen), as is the case with almost all passive pollinators. While senita moths, yucca moths, and fig wasps each have their own unique active pollination behaviors, these insects are the only three groups known in nature to exhibit active pollination behavior.

Following active pollination, female moths oviposit a single egg on the flower. In most cases, eggs are laid on the tips of flower petals, but they can also occur among anthers and within corolla tubes. By actively pollinating flowers, senita moths facilitate the survival of their offspring by increasing the likelihood of fruit set, as eggs and larvae in aborting flowers and fruit die. Before departing a flower, female moths actively collect pollen and in some cases nectar.

Within a few hours to 2-3 days after being laid, eggs hatch and larvae move down wilting corolla tubes toward the fruit's developing ovary. Larvae then bore into the fruit, consume developing ovules, and exit the base of the fruit and bore into the cactus stem where pupation occurs. This entire process, from oviposition to pupation, can be as short as one month. Not all eggs laid in flowers produce new adult moths, however, as many eggs and larvae die due to fruit abortions resulting from resource-limited fruit set in senita cacti.

Pollination by senita moths is typically sufficient enough that fruit set and seed/fruit production are resource-limited. While senita cacti rely almost entirely upon senita moths for reproduction, a few diurnal co-pollinators (halictid bees) do occur, but only very rarely do they make significant contributions to fruit set and plant reproduction.